




Orientador(a): Marcio Antonio Brunetto
Instituição: FMVZ USP
1º Colocado na 8ª Edição (2022) do Prêmio de Pesquisa PremieRpet®.
Índice
Resumo
A obesidade felina é uma afecção cada vez mais recorrente que pode implicar no desenvolvimento de diversas comorbidades, tais como resistência insulínica, diabetes e hiperlipidemia. Porém, pouco se sabe sobre o metaboloma destes animais, que por meio da correlação dos metabólitos alterados com rotas metabólicas pode possibilitar a obtenção de marcadores que alertem as comorbidades decorrentes da obesidade antes do surgimento dos sinais clínicos. Portanto, o objetivo desse estudo foi caracterizar e comparar o metaboloma sérico de gatos saudáveis e obesos. Foram incluídos 20 animais distribuídos em dois grupos de acordo com o escore de condição corporal (ECC), sendo o grupo controle “CO” composto por animais em ECC ideal (5/9) e o grupo obeso “OB” por animais com ECC indicativo de obesidade (8–9/9). Após triagem para verificar a higidez, os animais receberam uma dieta controle por 21 dias, a fim de padronizar a resposta nas análises. Após essa etapa os animais foram avaliados quanto a composição corporal pelo método de isótopos estáveis, análise de metabolômica sérica pelo método de cromatográfica gasosa acoplado a espectrometria de massas. Os dados obtidos foram submetidos a análise multivariada, onde foram realizados testes de robustez e reprodutibilidade pela técnica de PCA e análises de modelos supervisionados por PLS e OPLS, através da ferramenta Metaboanalyst. Também foi realizada análise estatística univariada dos dados para comparação entre os grupos e, foram considerados significativos valores de p<0,05. Os gatos obesos apresentaram maior quantidade de massa muscular (em kg), de massa gorda (em kg e %), ECC e peso corporal quando comparado ao grupo controle. No total, foram identificados 47 metabólitos e, os que apontaram diferença entre os grupos (p<0,05) foram valina, isoleucina, fenilalanina, tirosina, ácidos esteárico (AE), linoleico (AL), araquidônico (ARA) e alfa-tocoferol. Conclui-se que nas condições de realização do presente estudo, o metaboloma de gatos saudáveis difere do de gatos obesos. Os metabólitos encontrados nesse estudo destacam a influência de biomarcadores presentes na obesidade na evolução de outras comorbidades como resistência insulínica, diabetes e alterações do perfil lipídico.
Introdução
A obesidade é definida pela Organização Mundial da Saúde (OMS) como sendo o acúmulo anormal ou excessivo de gordura que pode prejudicar o bem-estar e a vida saudável. Em animais de companhia, esta condição é considerada presente quando o animal está 20% ou mais acima do seu peso corporal ideal (BURKHOLDER e TOLL, 2000). Esse acúmulo excessivo de gordura corporal pode resultar em comorbidades como resistência insulínica e diabetes em gatos (HOENIG et al., 2007). O status inflamatório ocasionado pela produção de adipocinas parece ter influência direta na origem destas alterações (VAN DE VELDE et al., 2013). Além disso, gatos obesos podem apresentar também dislipidemia, caracterizada por alterações no perfil de lipoproteínas e aumento de triglicerídeos e colesterol na circulação (HOENIG et al., 2013).
Estudos de metabolômica são comumente empregados na medicina humana afim de caracterizar e descobrir possíveis biomarcadores, demonstrar intervenção de dietas em rotas metabólicas específicas (BRENNAN, 2014), associar o tipo de dieta a doenças, ou a influência das dietas sobre uma determinada doença (CANUDO et al., 2018). Devido a abundância de informações, enorme diversidade de estruturas químicas e complexidade no estudo dos metabólitos, inúmeras informações podem ser obtidas através desta metodologia, porém poucos estudos buscaram avaliar o perfil metabolômico de gatos e nenhum deles com gatos obesos em comparação a gatos saudáveis. Portanto, o presente trabalho objetivou caracterizar e comparar o perfil de metabólitos séricos de gatos em condição corporal ideal e obesos.
Material e Métodos
Todo o protocolo experimental deste projeto foi conduzido de acordo com os princípios éticos de experimentação animal e, sob aprovação da Comissão de Ética no Uso de Animais (CEUA) nº 8713140219. Foram utilizados 20 gatos, os quais foram divididos em grupos de acordo com o escore de condição corporal (ECC), proposto por Laflamme (1997). O grupo controle (CO) foi constituído por 10 gatos com ECC ideal e o grupo obeso (OB) foi constituído por 10 animais com ECC 8 e 9. Os animais foram selecionados na rotina de uma clínica veterinária. Estes tiveram sua higidez avaliada previamente por meio de exame físico, hemograma e exames bioquímicos. Gatos com alterações laboratoriais ou sinais clínicos compatíveis com doenças endócrinas foram avaliados por meio de exames hormonais e, se positivos, não foram incluídos no estudo.
Posteriormente, os animais foram alimentados durante o período de 21 dias com alimento experimental de manutenção (Alimento Controle) com a finalidade de padronização, uma vez que todos os gatos eram domiciliados em lares diferentes. A equação utilizada para o cálculo da necessidade energética diária dos gatos em escore de condição ideal foi a de 100 x PC0,67 = kcal/ dia e, para os gatos obesos 130 x PC0,4= kcal/dia. A quantidade diária de alimento fornecida para cada animal foi determinada considerando-se a energia metabolizável do alimento.
Após a padronização da dieta foi determinada a composição corporal pelo método de diluição de isótopos de deutério (FERRIER et al., 2002; BRUNETTO et al., 2011). Para realização das coletas, os animais foram colocados em jejum alimentar prévio de 8 horas e hídrico por duas horas. Primeiramente, uma amostra de 2mL de sangue foi coletada através de punção da veia jugular. Posteriormente, foi inoculado 0,4g/Kg de 2H2O por via subcutânea. Passadas duas horas da inoculação, nova amostra de 2mL de sangue foi coletada e, na sequência foram processadas para extração de soro e então armazenadas a -20°C. Após a quantificação da água corpórea, foi calculada a massa magra total e por diferença a massa gorda (expressa na forma de porcentagem).
Para a avaliação de metabolômica, foram coletados 2mL de sangue por punção da veia jugular. As amostras foram centrifugadas por 15 min a 3000g e 4◦C; o soro sobrenadante foi transferido para tubos plásticos rotulados e armazenado a – 80°C, até a análise de cromatografia gasosa acoplada a espectrometria de massas (GCMS) através de metodologia descrita por Vallejo et al. (2009) e Naz et al. (2013).
Para a comparação da condição corporal entre os grupos foi realizado teste de ANOVA através do PROC MIXED do SAS. Já a análise dos perfis metabólitos foram verificados por análise univariada (ANOVA pelos pacotes do R) e multivariada pelo metaboanalyst. Valores de p<0,05 foram considerados significativos.
Resultados
Foram excluídos dois gatos de um mesmo tutor, um pertencente ao grupo obeso e o outro ao grupo controle por dificuldades de participação nas coletas experimentais. Ao final do projeto, 9 animais compuseram cada um dos dois grupos experimentais.
Os gatos obesos apresentaram maior quantidade de massa muscular (em kg), de massa gorda (em kg e %), ECC e peso corporal quando comparado ao grupo não obeso (Tabela 1; p<0,05).
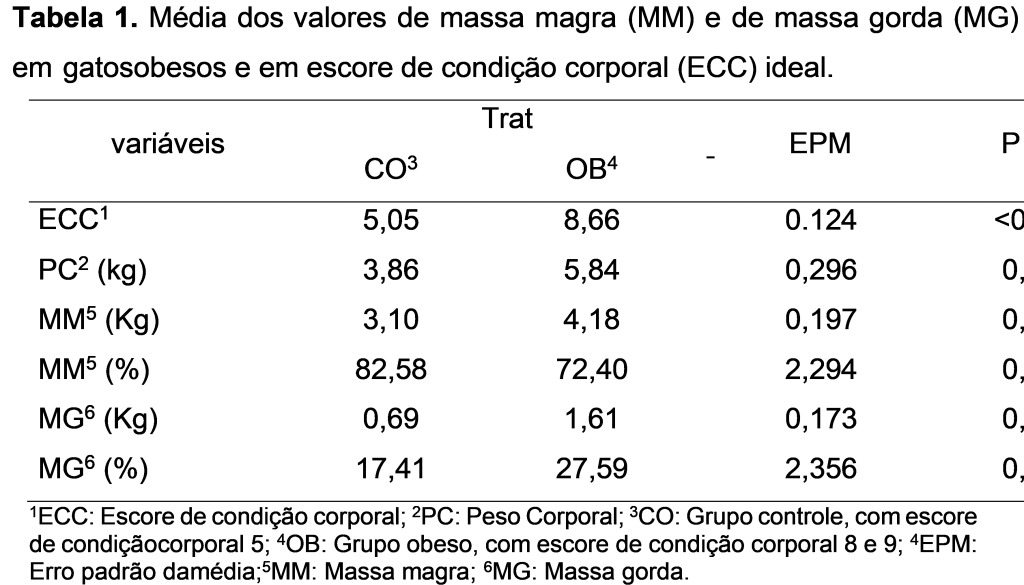
Foram identificados 47 metabólitos, porém aqueles com coeficiente de variação no controle de qualidade (QCs) acima de 30% para análises em GCMS foram removidos da lista do grupo de metabólitos que fizeram parte das análises estatísticas uni- e multivariada. Inicialmente os modelos testados de PLS para indivíduos tanto do grupo controle quanto do grupo de obesos não mostraram separação entre eles, apontando também variáveis preditivas e de validação cruzada muito baixas para os modelos propostos.
Devido à impossibilidade de evidenciar as diferenças metabólicas nos modelos multivariados analisados, foi realizado também teste estatístico de natureza univariada, utilizando o algoritmo estatístico de Análise de Variância com nível de significância de 5%. Os metabólitos que apontaram diferença entre os grupos (p<0,05) foram valina, isoleucina, fenilalanina, tirosina, ácidos esteárico (AE), linoleico (AL), araquidônico (ARA) e alfa-tocoferol, descritos na tabela 2.

Discussão
Os metabólitos são produtos intermediários ou finais do metabolismo em uma amostra biológica (FIEHN, 2002). O conjunto desses metabólitos que possuem massa molecular de até 1500 Da, presentes em um sistema biológico, é chamado de metaboloma (CANUDO et al., 2018). A metabolômica pode ser utilizada para dois objetivos: A análise de metabolômica alvo ou “targeted metabolomics”, definida como a análise quantitativa de um ou mais metabólitos pré-selecionados de determinada classe química, ou que estejam associados a rotas metabólicas específicas; e Análise de metabolômica global ou “untargeted metabolomics”, que está baseada na análise qualitativa do maior número de metabólitos possível, pertencentes a diversas classes químicas, contidas no sistema biológico avaliado (CANUDO et al., 2018). No presente estudo foi realizada análise de metabolomica untarget, com intuito de caracterizar o metaboloma desses grupos.
O estudo realizado por Pallotto e colaboradores (2021) avaliou gatos obesos durante o programa de perda de peso por um período de 16 semanas e, os autores observaram diferenças principalmente no metabolismo das gorduras nos animais, ao longo do emagrecimento, o que corrobora com os achados deste estudo, uma vez que ao se comparar os grupos foram observadas diferenças nas concentrações de AE, AL e ARA. Os autores sugeriram que provavelmente ocorreu diminuição desses metabólitos conforme os gatos do estudo foram perdendo peso.
Os lipídios presentes na corrente sanguínea podem ser derivados da dieta, do tecido adiposo ou do fígado. Esses lipídios são uma importante fonte de energia para o hospedeiro e são armazenados principalmente como triglicerídeos no tecido adiposo (PALLOTTO et al., 2021). A obesidade está geralmente associada a concentrações elevadas no plasma, soro e fígado de ácidos graxos não esterificados (WANG et al., 2003). Além disso, concentrações mais altas de AE associadas a concentrações mais baixas de ácido linoleico AL foram relatadas em homens obesos (KIM et al., 2010). Em humanos ocorrem várias alterações nos biomarcadores relacionados à obesidade, incluindo concentrações mais elevadas de insulina, glicose, índice HOMA e, concentrações mais baixas de adiponectina; e metabolismo anormal do ácido linoleico, como observado no presente estudo. Embora os gatos sejam considerados carnívoros e não desenvolvam doenças cardiovasculares da mesma forma que os humanos, o perfil lipídico verificado por meio do metaboloma sérico é um indicativo de seu estado metabólico (PALLOTTO et al., 2021).
O ARA é precursor de metabólitos eicosanóides, como prostaglandinas, tromboxanos, leucotrienos e lipoxinas (HAEGGSTRÖM et al., 2010). Eles são mediadores de inflamação aguda, febre e doenças como câncer, aterosclerose e trombose em humanos, comumente encontrados em pacientes com obesidade mórbida (LOPES & EGAN 2006).
Pallotto et al. (2021) observaram diferenças nos aminoácidos de cadeia ramificada (AACR) e aromáticos (AAA), que ao longo do emagrecimento foram apresentando diminuição em suas concentrações. Em humanos obesos as concentrações plasmáticas de valina e leucina são cerca de 23% e 14%, maior do que homens magros. O catabolismo de AACR pode ser inibido com a obesidade, isso já foi observado em camundongos e ratos obesos, os quais apresentaram diminuição na atividade da aminotransferase e do complexo enzimático α-cetoácido desidrogenase de cadeia ramificada (SHE et al., 2007).
Em estudo retrospectivo, concentrações elevadas de AACR foram relacionadas à idade avançada, sexo masculino, síndrome metabólica, obesidade, risco de doença cardiovascular, dislipidemia, hipertensão e ácido úrico (HU et al., 2016). Em humanos, a resistência à insulina e o risco aumentado ao desenvolvimento de diabetes mellitus tipo 2 foram associados a ingestão e concentrações elevadas de aminoácidos aromáticos (AAA) como tirosina e fenilalanina e AACR como isoleucina, leucina e valina, sendo relatado que os AA e AACR diminuem após a perda de peso em indivíduos obesos (SHE et al., 2007; WANG et al., 2011; WURTZ et al., 2012; NEWGARD, 2012).
As diferenças no perfil de metabólitos encontrados no presente estudo corroboram as informações publicadas em estudos com humanos, que já comprovaram a influência desses biomarcadores consequentes a obesidade no desenvolvimento de outras comorbidades: AAA e AACR na ocorrência de resistência insulínica e diabetes do tipo 2; e alterações do perfil lipídico, como aumento de AE e diminuição de AL em pessoas obesas.
Conclusões
Conclui-se que nas condições de realização do presente estudo, o metaboloma de gatos saudáveis difere do de gatos obesos. Os metabólitos que se diferiram destacam a influência de biomarcadores consequentes a obesidade na evolução de outras comorbidades como resistência insulínica, diabetes e alterações do perfil lipídico.
Referências bibliográficas
BRENNAN, C. L.; HOENIG, M.; FERGUSON, D. C. GLUT4 but not GLUT1 expression decreases early in the development of feline obesity. Domestic Animal Endocrinology. v. 26, n. 4, p. 291–301, 2004.
BRUNETTO, M. A. et al. Correspondência entre obesidade e hiperlipidemia em case. Ciencia Rural. v. 41, n. 2, p. 266–271, 2011.
BURKHOLDER, W. J.; TOLL, P. . Obesity. In: LEWIS, L. D.; MORRIS, M. L.; HAND,. (Ed.). Small Animal Clinical Nutrition. 4th. ed. [s.l.] Topeka, Kansas: Mark Morris Institut, p. 401–425, 2000.
CANUTO, Gisele AB et al. Metabolômica: definições, estado-da-arte e aplicações representativas. Química Nova, v. 41, p. 75-91, 2018.
FERRIER, L. et al. Evaluation of Body Composition in Dogs by Isotopic Dilution Using a Low-Cost Technique, Fourier-Transform Infrared Spectroscopy. The Journal of Nutrition. v. 132, n. 6, p. 1725S-1727S, 2002.
FIEHN, Oliver. Metabolomics—the link between genotypes and phenotypes. Functional genomics, p. 155-171, 2002.
Haeggström J.Z., Rinaldo-Matthis A., Wheelock C.E., Wetterholm A. Advances in eicosanoid research, novel therapeutic implications. Biochemical and Biophysical Research Communications. v. 396, 135–139, 2010.
HOENIG, M. et al. Cats differ from other species in their cytokine and antioxidant enzyme response when developing obesity. Obesity, v. 21, n. 9, p. 407–414, 2013.
HOENIG, M. et al. Insulin sensitivity, fat distribution, and adipocytokine response to different diets in lean and obese cats before and after weight loss. American Journal of Physiology – Regulatory Integrative and Comparative Physiology, v. 292, n. 1, p. 227–234, 2007.
HU, Wen et al. Relationship between branched-chain amino acids, metabolic syndrome, and cardiovascular risk profile in a Chinese population: a cross-sectional study. International journal of endocrinology, v. 2016, 2016.
KIM, Ji Young et al. Metabolic profiling of plasma in overweight/obese and lean men using ultra performance liquid chromatography and Q-TOF mass spectrometry (UPLC− Q-TOF MS). Journal of proteome research, v. 9, n. 9, p. 4368-4375, 2010.
LAFLAMME, D. R. P. C. Development and validation of a body condition score system for cats: a clinical tool. Feline Practice, v. 25, p. 13–18, 1997.
LOPES, Heno Ferreira; EGAN, Brent M. Desequilíbrio autonômico e síndrome metabólica: parceiros patológicos em uma pandemia global emergente. Arquivos Brasileiros de Cardiologia, v. 87, n. 4, p. 538-547, 2006.
NAZ, Shama; GARCÍA, Antonia; BARBAS, Coral. Multiplatform analytical methodology for metabolic fingerprinting of lung tissue. Analytical chemistry, v. 85, n. 22, p. 10941- 10948, 2013.
PALLOTTO, Marissa R. et al. Effects of Weight Loss and Moderate-Protein, High-Fiber Diet Consumption on the Fasted Serum Metabolome of Cats. Metabolites, v. 11, n. 5, p. 324, 2021.
SHE, Pengxiang et al. Obesity-related elevations in plasma leucine are associated with alterations in enzymes involved in branched-chain amino acid metabolism. American Journal of Physiology-Endocrinology and Metabolism, v. 293, n. 6, p. E1552- E1563, 2007.
VALLEJO, M. et al. Plasma fingerprinting with GC-MS in acute coronary syndrome. Analytical and Bioanalytical Chemistry, v. 394, n. 6, p. 1517-1524, 2009.
VAN DE VELDE, H. et al. The cat as a model for human obesity: Insights into depot- specific inflammation associated with feline obesity. British Journal of Nutrition, v. 110, n. 7, p. 1326–1335, 2013.
WANG LU, et al., Plasma fatty acid composition and incidence of diabetes in middle- aged adults: the Atherosclerosis Risk in Communities (ARIC) Study. The American journal of clinical nutrition, v. 78, n. 1, p. 91-98, 2003.
WANG, Thomas J. et al. Metabolite profiles and the risk of developing diabetes. Nature medicine, v. 17, n. 4, p. 448-453, 2011.
WÜRTZ, Peter et al. Metabolic signatures of insulin resistance in 7,098 young adults. Diabetes, v. 61, n. 6, p. 1372-1380, 2012.
NEWGARD, Christopher B. Interplay between lipids and branched-chain amino acids in development of insulin resistance. Cell metabolism, v. 15, n. 5, p. 606-614, 2012.


